Brown2 project protocol
Visual ability and spatial, motor and olfactory learning and memory in 14 inbred strains of mice (2004)
Brown RE, Schellinck HM, Gunn RK, Wong AA, O'Leary TPWith: Currie L, Hoffman N, Savoie V
Project protocol — Contents
Definitions
Overall workflow
Equipment common to multiple procedures
Notes for all procedures in this project
Procedure: Barnes Circular Maze (BCM) for testing visuo-spatial learning and memory
Procedure: Rotarod (ROD) for measuring neuromuscular coordination, cerebellar function, and motor learning
Procedure: Olfactory discrimination task (ODT) to measure olfactory discrimination learning and memory
Procedure: Morris Water Maze (MWM)
Procedure: Visual Water Box (VWB) to assesses visual discrimination, pattern discrimination and visual acuity
Data
ReferencesThe majority, but not all mice were tested for all procedures. Mice were not pretreated with drugs, special diet, or exposed to other environmental test conditions.
Week
Test
2-3*
4-5
5-6
7-8
9-14
* Week-1 is devoted to testing anxiety and exploratory behavior (Brown1).
Equipment common to multiple procedures
- Camera-based computer tracking system: Limelight and Watermaze, Actimetrics; IBM PC computer) fixed to the ceiling 2.1 m above the apparatus for measuring latency and distance traveled.
- Video camcorder: (Hitachi, VM-7500LA) located 150 cm above the apparatus for recording trials for subsequent analysis (with Hindsight).
- Hindsight event recording software: (MS-dos, version 1.5, Scott Weiss) for scoring behavior.
Notes for all procedures in this project
- Mice are transported to the test room in their home cage.
- Mice are handled by the base of the tail or with a 500 ml plastic container when moved in and out of the apparatus
- An observer records the behavior of each mouse from a distance of 1 m.
- Each trial is recorded with a video camera positioned above the apparatus (for subsequent computer analysis).
- The apparatus is thoroughly cleaned with 70% EtOH and allowed to dry between tests.
----------------------------------
Purpose: To measure strain differences in visuo-spatial learning and memory in 14 inbred strains of mice.
Workflow and Sampling
Workflow
Habituation Bright lights Acquisition training Bright lights, buzzer Reversal training Bright lights, buzzer Probe trial Bright lights Equipment
Barnes Circular Maze: The Barnes maze was originally developed by Carol Barnes as a method for testing visuo-spatial learning in aging rats (Barnes 1979; Barnes, Nadel, & Honig, 1980). The Barnes Maze for mice used here is a modified version of that developed by Pompl et al. (1999). In this maze mice are motivated by aversive bright light and loud noise to locate a hole which leads to a dark escape box beneath the maze.
The Barnes Maze was a 69 cm diameter circular disk constructed from 2 cm thick plywood and painted white with non-toxic oil-based paint. A 15 cm high wall made from white polyethylene was placed around the edge of the disk. There were 16, 4.45 cm diameter circular holes equally spaced around the edge of the maze and these holes were located 1.3 cm from the wall.
The circular disk was placed on top of a circular platform that was 89 cm in diameter and 1 cm thick. The platform had four legs screwed into the bottom that raised the maze 48.4 cm above the floor. The holes on the circular disk went through this platform and these holes were labeled from 1 to 16 on the outside of the wall.
An escape box (13 x 29 x 14 cm) made of white plastic could be placed under four different escape holes (holes 4, 8, 12 and 16). A staircase made of plywood was placed in the escape box to ease the descent of mice from the test arena into the escape box. This staircase had five stairs (each 1.8 cm high, 1.3 cm long and 10 cm wide) and the top of the stairs was 4 cm below the escape hole. The bottom of the staircase was 18 cm long and 10 cm wide, while the top of the staircase was 7.7 cm long and 10 cm wide. The entire set of stairs was 9.0 cm high. The stairs rested on a piece of 1 cm thick plywood in the bottom of the plastic escape box. The top of the staircase was painted white, while the rest of the stairs were painted black.
A blue polyvinyl chloride (PVC) tube that was 8 cm in diameter and 12.5 cm high was placed in center of the maze floor and mice were placed into this tube before each trial.
A light stand held two 150-Watt light bulbs (emitted 2580 lux) 76 cm above the maze platform to provide aversive bright light. Also a buzzer powered by a 9-volt battery was attached to the light stand and hung 20 cm above the center of the maze. The buzzer emitted a 372 kHz sound at 89 decibels directly under the buzzer (86 decibels at the wall). The buzzer was turned on and off by the experimenter using a push button.

Figure 1 is a schematic illustration of the Barnes circular maze apparatus.
Visual cues: Three intra-maze visual cues (geometric shapes) were taped to the inside of the wall in semi-random locations, so that no cue was located directly above an escape hole. Also objects around the room (room walls, shelves, desk, computer, light stand etc.) served as extra-maze visual cues.
Environmental test conditions
The Barnes maze was located in the same laboratory room (1.8 x 4.5 m) where testing on the elevated zero maze, elevated plus maze, light-dark box and open field test previously occurred. This room had a video camcorder mounted on a bracket 150 cm above the center of the maze to videotape test trials for later analysis. The room was kept dark except for the bright lights above of the Barnes maze.
Procedure: Barnes Circular Maze (BCM)
See details common to all procedures in this project, and see Table 1 Barnes maze for a greater detail of the test.
Before testing, mice are randomly assigned to one of four different escape hole locations, which determined where the escape hole would be located throughout testing on the Barnes maze.
Mice first completed habituation training (Day 1), which consisted of four trials. In each trial mice are placed next to their escape hole, under an inverted 2 L glass beaker. Following habituation training, mice completed acquisition training for 4 days (4 trials per day). On each trial, mice are released from the start tube and are given 5 minutes to enter the escape hole. Following acquisition training, mice completed 4 days of reversal training where the escape hole is moved to the opposite side of the maze.
During the probe trial no escape box is present and the trial is videotaped and later analyzed using the Hindsight event recorder program. The maze is divided into four quadrants, and the time spent in each quadrant is measured. Memory for the location of the escape hole is assessed with the % time in the quadrant where the escape hole is located in previous training.
Data collected by investigator
The numbers of errors made (head-dips into incorrect holes) and latency to find the escape hole were recorded on each trial of acquisition and reversal training to assess learning. Also, distance traveled during each trial on the last day (day 4) of reversal training was used to assess learning. Entries and time spent in the correct, right, left, and opposite quadrants were recorded during the probe trial to assess memory. Also the number of nose pokes into the acquisition and reversal escape holes during the probe trial were recorded to assess memory.
Distance traveled was obtained with analysis of the video recorded trials, by placing transparent plastic sheets over a television screen, and tracing the movement of each mouse with a marker. The distance traveled and the diameter of the maze on the plastic transparencies (Scalex, PlanWheel XL) is then used to determine the distance traveled in the actual maze using the formula [(Real Maze Diameter ÷ Transparent Sheet Maze Diameter) X Tracing Distance]. Distance traveled can also be obtained using a video-camera based tracking system such as Limelight (Actimetrics).
Investigator's notes: The Barnes maze was originally designed for rats and the original apparatus design is often altered when the maze is used with mice. For example, the maze used here has a smaller size (69 cm diameter) and has fewer holes (16) than the original design for rats (122 cm diameter and 18 holes). Also, a wall and intra-maze cues have been added around the edge of the maze. Other designs have been used with mice that have more holes (40) (Bach et al., 1995; Fox et al., 1998), or use different stimuli to provide aversive motivation (tactile fan or auditory buzzer) (Pompl et al., 1999). Little research has been completed to determine if results using one Barnes maze design can generalize to other Barnes maze designs. Therefore, differences in apparatus design should be considered when comparing results from previous studies to the present database.
The validity of the Barnes maze design used in the present database has been questioned. We found that C57BL/6J mice on this design may not use visual cues to locate the escape hole as performance of mice did not change when extra-maze visual cues were not visible, or when the location of the escape hole was moved. Also, Garcia et al. (2004) found that the performance of blind mice did not differ from sighted mice on this design. Therefore, results from this Barnes maze design should be considered in conjunction with results from another visuo-spatial learning and memory test (i.e. Morris water maze).
---------------------------------------------
Purpose: To measure strain differences in neuromuscular coordination and motor learning using the rotarod apparatus.
Equipment
Rotarod: The rotarod apparatus (AccuRotor Rotarod; Accuscan Instruments Inc. Columbus, Ohio (number: AI9612AR04/M)) was capable of accommodating four mice at the same time, with each mouse separated by a 15 cm high white Plexiglas circular divider. The space between each divider was 11 cm. The rotating rod was acrylic and measured 3 cm in thickness. There was a drop of 39 cm from the rod to the individual holding chambers that were directly below each section of the rotating rod.
Four automatic timers were built into the apparatus and these timers started the moment the rod was set into motion at the start of each trial, and turned off automatically when an animal fell into the holding chamber below the rod.

Figure 2 is an example of a Rotarod apparatus. Note the mice positioned in different trails.
Environmental test conditions
The Rotarod was located in a laboratory room measuring 112 cm x 260 cm. The room was illuminated with a single 60-Watt red light bulb.
Procedure: Rotarod
Data collected by investigatora. See details common to all procedures in this project, Table 2 Rotarod, and Brown and Wong (2007) for greater details of this test.
b. Mice are tested in squads of 4 and are tested during the dark phase of the Light/Dark cycle.
c. For each trial, four mice are placed on the rod facing the direction opposite to the rod's motion. The speed of the rod increased from 0 rotation per min (rpm) to a maximum speed of 48 rpm over each 6 min trial.
d. Between trials mice are given 1 min to rest in the holding chambers until the next trial.
e. To measure motor learning, mice completed six trials per day for seven days.Days 1-7 of sensorimotor learning: the latency for mice to fall off the accelerating-rotating rod, and body weight data were recorded.
Investigator notes: The Rotarod is a measure of neuromuscular co-ordination, fatigue, and learning (LeMarec & Lalonde, 1997). Rotarod (or Roto rod) performance has often been used as a method to study the effects of drugs on neuromuscular co-ordination (Hymson & Hynes, 1982), a measure of cerebellar function (Caston et al., 1998) and a measure of motor learning (Hyde et al., 2001). The Rotarod can rotate at a constant speed or accelerate over time. The accelerating Rotarod is better at detecting drug effects and a better model of motor learning than the constant speed Rotarod (Bogo et al., 1981). The Procedure as described here is a sensorimotor learning task. The latency for the mice to fall off the rod will increase over trials and over days as the animal becomes more proficient with maneuvering on the apparatus and can stay on longer.
Rotarod performance is sensitive to body weight with mice of smaller strains having longer latencies to fall then mice of larger strains and females having a longer latency then males. Thus it is important to record body weights and to use an analysis of co-variance with weight as the co-variate to examine strain x sex differences (McFadyen et al., 2003; Brown, unpublished).
-------------------------------------
Olfactory discrimination task (ODT)
Purpose: To measure olfactory discrimination learning and memory by using a classical conditioning paradigm.
Workflow and Sampling
Workflow
Food restriction bw Training bw Odor discrimination bw, time spent (L or R) Odor discrimination bw, time spent digging Equipment
The odor discrimination test apparatus consisted of a 69 x 20 x 20 cm box made of 3 mm acrylic and was divided into three compartments of equal size (23 x 20 x 20 cm) by two walls. These walls had 6 x 5.5 cm openings located at floor level, and allowed mice to move between compartments of the apparatus.
Before each trial, the floor of the test apparatus was covered with 1000 ml of Pro-chip (P.W. I Industries) and acrylic doors were placed over the openings in the walls between the chambers. These doors were removed at the start of each test trial.
Odor training apparatus: Training trials were completed in polycarbonate cages with stainless steel wire tops, identical to the cages mice were housed in.

Figure 3 illustrates the typical layout of an odor discrimination preference apparatus.
Odor pot: A diagram of the odor pot used to present the odors during training and testing (From Schellink 2001 a) is shown below. Odor pots contained rose or lemon scent (15%, 3 ml of phenyl acetate (rose) or linalool (lemon) in 17 ml of propylene glycol), 6-8 small pieces of sugar (cut from sugar cubes), and pine chip bedding.
Odors were stored in 1.5 ml Eppendorf tubes at –80°C to avoid degradation.
The odor pots were covered with perforated Petri dishes to prevent physical contact with odorants, while allowing mice to smell the odors.
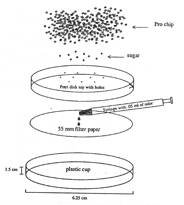
Figure 4 is a schematic diagram of the odor pot setup from Schellink 2001 a.
- Pots: trimmed plastic beverage cups (1.5 cm in height and 6.25 cm diameter).
- Odors: Phenyl acetate (Rose) and Linalool (Lemon) (Aldrich Chemicals).
- Diluent: propylene glycol (Caledon Chemical Co.).
- Adsorbent paper: Whatman® no.1 filter paper (55 mm diameter).
- Reinforcement: sugar cubes pre-cut with razor blade.
- Digging medium: pine chips (Pro-chip, P.W. I. Industries) bedding.
Procedure: Olfactory discrimination task (ODT)
See details common to all procedures in this project, Table 3 olfactory discrimination, and see Schellinck et al (2001a & 2001b) for more in-depth details of the task.
a. Three days before training mice are placed under food restriction until each mouse's body weight is reduced to 85-90% of its body weight when food is available ad libitum. During food restriction mice are weighed and fed more or less than 3g of food to maintain their body weight at 85-90% of their ad-libitum weight. Mice are fed shortly after lights off (9:00 am). Food restriction is continued throughout training. After the completion of the discrimination test (day 8), food is again made available ad libitum.
b. Mice are weighed each day before training trials, and fed 3 hours after each day's training is finished (~4:00 pm).
c. Training occurred in three separate rooms, one for the rose odor, one for the lemon odor, and one neutral. Mice are kept in the neutral room during inter-trial intervals.
d. Half of the mice are trained with the rose odor paired with sugar (CS+) and the lemon odor not paired with sugar (CS-). The other half of the mice are trained with the lemon odor paired with sugar (CS+) and the rose odor not paired with sugar (CS-). In each training trial an odor pot is placed in the center of the back third of the training cage. Each mouse received four 10-min training trials per day on each of 4 days. Two trials are completed with the rose odor and two trials are completed with the lemon odor and these trials are presented in a pseudo-random order.
e. The odor discrimination test is done in a novel room to avoid any influence of contextual cues from rooms used during odor training.
f. During the habituation trial a pot (no odors or sugar) is placed in each of the end compartments. Each mouse is placed in the center compartment and are allowed to explore the apparatus for 3 min.
g. To prevent the use of visual cues, the apparatus is rotated 180 degrees for the preference test. Odor pots (without sugar) containing rose and lemon odors are placed in each of the end compartments. During the discrimination trial each mouse is placed in the center chamber, and is allowed to explore the apparatus for 3 min.
h. The time spent digging in the rose and lemon odor pots is measured to assess memory for the odor that is paired with sugar. This test phase is sensitive to the level of food restriction in mice (Forestell et al., 2001).
i. Bedding and odor pots are replaced between test trials, and the odor discrimination apparatus is cleaned between trials.Data collected by investigator
In addition to behavioral measures: body weight on each day of training, body weight before training began with food ad libitum, body weight on the day of odor discrimination testing.
During the habituation trial time spent in each compartment was measured to determine pre-existing preferences for different compartments of the apparatus.
Memory for odor-sugar pairing was measured in the odor discrimination test with time spent digging at the Cs+ odor pot, time spent digging at the Cs- odor pot, and percentage of the total time digging that each mouse dug at the Cs+ odor pot.
Investigator notes: Mice were placed under a food restricted diet so that they are motivated to obtain sugar during training and during the odor preference test. If mice are not food deprived during the odor preference test, they do not accurately discriminate between the two odors (Forestall et al., 2001). Therefore, the body weight of mice on the preference test should be considered when interpreting the percent time digging in the Cs+ odor pot.
------------------------------------------
Purpose: To test visuo-spatial learning and memory on the Morris Water Maze.
Workflow and Sampling
Workflow
Acquisition Invisible, north west Reversal Invisible, south east Probe none Visible platform Visible, south west Equipment
- The Morris water maze (MWM) was originally designed to assess visuo-spatial learning and memory in rats (Morris, 1984). A modified version of the MWM for mice (Paylor et al., 1996) was used and consisted of a circular polypropylene pool (Canadian Tire "Pelican" pool) that was 110 cm in diameter and 20 cm in height. The pool was filled with water to a dept of 14 cm, and was kept at a 20±1°C temperature.
- The water was made opaque with 500 ml of non-toxic white liquid tempura paint (Schola, Marieville) to camouflage the white escape platform.
- A Plexiglas cylinder (13.75 cm x 9 cm diameter) was used as the escape platform in the maze. The cylinder was filled with water to weigh it down in the pool. The escape platform sat 0.5 cm below the surface of the water. A removable red and yellow striped top (3 cm x 9 cm in diameter) with a colorful flag erected in the center could be placed on the top of the escape platform to make the escape platform visible in the water.
- A video camera-based computer (IBM PC) tracking system (WaterMaze, Actimetrics) mounted to the ceiling 2.1 m above the pool, was used to record the mice water maze performance.
- Visual cues: Extra-maze visual cues around the maze consisted of a table, computer, and posters placed along the walls of the test room.
- The pool is divided into four quadrants: Northwest, Northeast, Southwest and Southeast. Boundaries of these quadrants are marked on the edges of the pool with masking tape and labeled: North, South, East and West.
Figure 5 is a photographic image of a Morris water maze.Environmental test conditions
The pool is located in a room measuring 5.2 x 2.4 m, and the test room was diffusely lit with white light (30 lux).
Procedure: Morris Water Maze (MWM)
See details common to all procedures in this project, and see Table 4A Morris water maze and Brown and Wong (2007) for specifics.
a. Mice are tested in squads of 4 and each mouse is placed in a clean empty cage during testing. Paper towel is torn and placed in the bottom of the cage to allow mice to dry quickly between trials. The paper towel is replaced when it became completely wet.
b. Mice completed 3 days of acquisition training with 4 trials per day. During each trial the mice are placed in the pool at one of four different start locations (North, South, East and West), and are given 60 s to find the hidden escape platform. To prevent mice from developing fixed motor patterns, the start locations varied across trials based on a latin square design (see Table 4B Morris water maze).
c. Following acquisition training, mice completed 3 days of reversal training (4 trials per day), where the escape platform is moved to the opposite side of the maze.
d. During the probe trial (day 8), the escape platform is not present. The maze is divided into 4 quadrants using the Watermaze (Actimetrics) software, and the time spent in each quadrant is recorded.
e. The day following the probe trial, mice completed visible platform training where a striped top was placed on top of the escape platform so that the escape platform was visible. Mice completed one day of visible platform training with four trials.Data collected by investigator
During acquisition and reversal training learning was assessed with the measures of swim latency (time to climb onto the escape platform) and distance traveled to reach the escape platform. Average velocity and the duration and frequency of thigmotaxic behavior (9-cm-wide corridor) were also recorded.
During the probe trial time spent in each quadrant (Northeast, Southeast, Northwest, Southwest) was recorded and memory was assessed with time spent in the quadrant that contained the escape platform during reversal training (southeast or correct quadrant). Memory was also assessed with the number of times each mouse crossed the location where the escape platform was located during reversal training (annulus reversal crossing) or acquisition training (annulus acquisition crossing).
Overall distance traveled is determined by the formula: [Real Maze Diameter/ Transparent Sheet Maze Diameter) X Tracing Distance].
Investigator notes: The swim path analysis can be used to determine whether mice are developing a spatial strategy for learning the maze or are using non-spatial strategies, such as circular swimming or thigmotaxis. Analysis of swim speed may be necessary to control for locomotor differences between strains (Wolfer et al., 1998).
Wolfer et al., 1998 found 3 factors which accounted for 81% of the observed variability in the scores of mice in the Morris water maze: thigmotaxis (49%), passivity (19%) and memory (13%). Thigmotaxis is associated with swimming along the wall of the pool; while passivity means floating in the pool, with a slow swimming speed; and memory represents the time spent in the target quadrant during the probe trial.

----------------------------
Purpose: To test visual discrimination, pattern discrimination and visual acuity in mice.
Workflow and Sampling
Mice are first gradually trained to locate a hidden platform below a screen displaying a vertical grating of low spatial frequency (S+) (see Wong & Brown, 2006).
Workflow
Pre-training Vertical, 0.17 (c/deg) Visual detection Vertical, 0.17 (c/deg) Pattern discrimination Vertical*, horizontal, 0.17 (c/deg) Visual acuity Vertical, 0.17 to 0.64 (c/deg) *Only the vertical grating is paired with the hidden platform.
Equipment
The visual water box combined the basic principles of a discrimination box (Yerkes, 1907) and the Morris water maze (Morris, 1984). The apparatus consisted of a trapezoidal-shaped pool made from 6 mm clear Plexiglas, and had 55 cm high walls around the edge. The walls on the smallest and longest sides of the pool were painted black to reduce reflections. The wall at the wide edge of the pool remained clear, so that two adjacent computer monitors placed behind this wall were visible.
A black Plexiglas divider (41 x 40 cm) was placed in guides between the computer screens, and extended from the clear wall, bisecting the end of the pool along its midline.
A black release alley (35 cm long x 7 cm wide x 20 cm high) was placed at the narrow end of the pool.
A movable clear Plexiglas escape platform (37cm long x 13cm wide x 14cm high) was placed below the computer screen displaying the positive (S+) visual stimulus. The pool and computer monitors were placed on a solid table (146 cm long x 100 cm wide x 46 cm high).
The pool was filled with tepid (22 °C) water to a depth of 15 cm. Reflections from the computer screens on the surface of the water made the platform invisible from water level.
Grating setup: Gratings were displayed on two identical 17-inch monitors (Accu Sync 70 NEC 3) that face into the wide end of the pool, and the bottom of the monitors were at water level.
A customized computer program (developed by Stephen Turner and modeled after the Vista program used by Prusky et al, (2000)) controlled the visual stimuli.

Figure 6 depicts the schematic layout of the visual water box. Note that lines are drawn for illustrative purposes.
Environmental test conditions
Testing was completed during the active (dark) phase of the Light/Dark cycle without any other lights present, except that from the computer screen.
Procedure: Visual Water Box (VWB)
See details common to all procedures in this project, and see Table 5 visual water box, and Wong & Brown, 2007 for specific details of the test.
a. Mice are trained in groups of 4 and given an inter-trial interval of about 3 min. Mice completed 4 phases of testing: pre-training, visual discrimination, pattern discrimination and visual acuity.
b. During pre-training (day 1), mice are shaped to locate a hidden platform situated below a computer monitor displaying a low spatial frequency vertical grating. The positive stimulus (S+, hidden platform) alternated between the left and the right side of the water box divider during testing. The negative stimulus (S-) is a grey screen and no escape platform is placed beneath it.
c. Mice that did not find the escape platform within 1 min are guided to the escape platform. Mice are allowed to stay on the platform for 10 s before being returned to the holding cages.
d. Visual discrimination testing (days 2-9) is completed following pre-training, and consisted of 8 days of training (8 trials/ day). During visual discrimination the escape platform is located under the monitor displaying vertical gratings (0.17 c/deg) (S+), whereas no escape platform is located under the monitor displaying a grey screen (S-). A trial is correct if an animal swam to the escape platform without entering the negative stimulus arm. If an animal broke the plane perpendicular to the end of the divider on the side of the tank with the monitor displaying the S- stimulus, the trial is recorded as an error. A trial is also incorrect if mice did not find the escape platform in less than 1 min.
e. If a mouse made an error, the mouse is required to run another trial (error trial) before being returned to its holding cage. Only one error trial is completed for each training trial.
f. Pattern discrimination training (days 10-17) is completed following visual discrimination training and consisted of 8 days of training (8 trials/day). During pattern discrimination the escape platform is located under a monitor displaying vertical gratings (S+) (0.17 c/deg), whereas no escape platform is located under a monitor displaying horizontal gratings (S-).
g. Visual acuity testing (days 18-25) is completed following pattern discrimination testing, and consisted of 8 days of testing (8 trials/day). On each day the escape platform is located under the monitor displaying vertical gratings (S+), whereas no escape platform was located under the monitor displaying a grey screen (S-). The spatial frequency of the vertical gratings ranged from 0.17 to 0.64 c/deg across trials for each day of testing.
h. Testing is accomplished efficiently and timely to prevent hypothermia and exhaustion in the mice.Data collected by investigator
For visual discrimination and pattern discrimination the percentage of correct trials for each day (8 trials per day) and latency to reach the platform were recorded as measures of learning. Data from error trials were not used. If a mouse performed above 70% (~6/8) correct response on two consecutive days, it met the criteria for that phase.
For visual acuity, the percent correct at each spatial frequency was recorded. The visual acuity was defined as the highest spatial frequency at which mice made more than 70% correct choices (~6/8 trials). Latency to find the escape platform was also recorded for each spatial frequency.
Investigator notes:Prolonged testing does not guarantee accurate results, because animals can get hypothermic and tired. Therefore, the holding cages of the mice were placed under a standard 60W light bulb as a source of heat and the holding cages were lined with paper towel to dry mice between trials. However, mice appeared to be visibly cold or tired, they were allowed to rest before continuing the experiment.
----------------------------------
Investigator's suggested reading list
- Akirav, I., Sandi, C. & Richter-Levin, G. (2001). Differential activation of hippocampus and amygdala following spatial learning under stress. European Journal of Neuroscience, 14, 719-725. Pubmed: 11556896
- Caston, J., Devulder, B., Jouen, F., Lalonde, R. (1999). Role of an enriched environment on the restoration of behavioral deficits in Lurcher mutant mice. Developmental Psychobioloy, 35, 291-303. Pubmed: 10573569
- Chapillon, P. & Debouzie, A. (2000). BALB/c mice are not so bad in the Morris water maze. Behavioral Brain Research, 117, 115-118. Pubmed: 11099764
- Chen, G., Chen, K.S., Knox, J., Inglis, J., Bernard, A., Martin, S.J, Justice, A., McConlogue, L., Games, D., Freedman, S.B. & Morris, R. (2000). A learning deficit related to age and beta-amyloid plaques in a mouse model of Alzheimer's disease. Nature, 408, 975-979. Pubmed: 11140684
- Dalm, S., Grootendorst, J., Kloet, E. R. and Oitzl, M. S. (2000). Quantification of swim patterns in the Morris water maze. Behavior Research Methods, Instruments & Computers, 32, 134-139. Pubmed: 10758671
- D'Hooge, R. & De Deyn, P., P. (2001). Applications of the Morris water maze in the study of learning and memory. Brain Research Reviews, 36, 60 – 90. Pubmed: 11516773
- Fellows, B. J. (1967) Chance stimulus sequences for discrimination tasks. Psychological Bulletin, 67, (87-92). Pubmed: 6045339
- Fordyce, D., Clark, V., Paylor, R. & Wehner, J. (1995). Enhancement of hippocampally-mediated learning and protein kinase C activity by oxiracetam in learning-impaired DBA/2 mice. Brain Research, 672, 170-176. Pubmed: 7749739
- Frick, K.M., Stillner, E.T., & Berger-Sweeney, J. (2000). Mice are not little rats: species differences in a one-day water maze task. Neuroreport, 11, 3461-3465. Pubmed: 11095500
- Gallagher, M., Burwell, R. and Burchinal, M. (1993). Severity of spatial learning impairment in aging: Development of a learning index for performance in the Morris water maze. Behavioral Neuroscience, 107: 618-626. Pubmed: 8397866
- Klapdor, K. & Van Der Staay, F.J. (1996). The Morris water-escape task in mice: strain differences and effects of intra-maze contrast and brightness. Physiology of Behavior, 60, 1247-1254. Pubmed: 8916178
- McDonald, R.J. & White, N.M. (1994). Parallel information processing in the water maze: evidence for independent memory systems involving dorsal striatum and hippocampus. Behavioral Neural Biology, 61, 260-270. Pubmed: 8067981
- Owen, E.H., Logue, S.F., Rasmussen, D.L. & Wehner, J. (1997). Assessment of learning by the Morris water task and fear conditioning in inbred mouse strains and F1 hybrids: implications of genetic backgrounds for single gene mutations and quantitative trait loci analyses. Neuroscience, 80, 1087-1099. Pubmed: 9284062
- Paradee, W., Melikian, H.E., Rasmussen, D.L., Kenneson, A., Conn, P.J. & Warren, S.T. (1999). Fragile X mouse: strain effects of knockout phenotype and evidence suggesting deficient amygdala function. Neuroscience, 94, 185-192. Pubmed: 10613508
- Spooner, R.I., Thomson, A., Hall, J., Morris, R. & Salter, S. (1994). The Atlantis platform: a new design and further developments of Buresova's on-demand platform for the water maze. Learning & Memory, 1, 203-211. Pubmed: 10467597
- Upchurch, M. & Wehner, J.M. (1988). Differences between inbred strains of mice in Morris water maze performance. Behavioral Genetics, 18, 55-68. Pubmed: 3365197
- Von Lubitz, D. A., Paul, I. A., Bartus, R. T. & Jacobson, K. A. (1993). Effects of chronic administration of adenosine A1 receptor agonist and antagonist on spatial learning and memory. European Journal of Pharmacology, 249, 271-280. Pubmed: 8287914
- Wishaw, I. A. & Jarrard, L. E. (1995). Similarities vs. differences in place learning and circadian activity in rats after fimbria-fornix section or ibotenate removal of hippocampal cells. Hippocampus 5, 595-604. Pubmed: 8646285
- Whishaw, I.Q. (1985). Cholinergic receptor blockade in the rat impairs local but not taxon strategies for place navigation in a swimming pool. Behavioral Neuroscience, 99: 979-1005. Pubmed: 3843314
References
Bach ME, Hawkins RD, Osman M, Kandel ER, Mayford M. Impairment of spatial but not contextual memory in CaMKII mutant mice with a selective loss of hippocampal LTP in the range of the theta frequency. Cell. 1995 Jun 16;81(6):905-15.
PubMed 7781067 Barnes CA. Memory deficits associated with senescence: a neurophysiological and behavioral study in the rat. J Comp Physiol Psychol. 1979 Feb;93(1):74-104.
PubMed 221551 Barnes CA, Nadel L, Honig WK. Spatial memory deficit in senescent rats. Can J Psychol. 1980 Mar;34(1):29-39.
PubMed 7388694 Bogo V, Hill TA, Young RW. Comparison of accelerod and rotarod sensitivity in detecting ethanol- and acrylamide-induced performance decrement in rats: review of ex perimental considerations of rotating rod systems. Neurotoxicology. 1981 Dec;2(4):765-87.
PubMed 7200586 Forestell CA, Schellinck HM, Boudreau SE, LoLordo VM. Effect of food restriction on acquisition and expression of a conditioned odor discrimination in mice. Physio l Behav. 2001 Mar;72(4):559-66.
PubMed 11282140 Fox GB, Fan L, LeVasseur RA, Faden AI. Effect of traumatic brain injury on mouse spatial and nonspatial learning in the Barnes circular maze. J Neurotrauma. 1998 D ec;15(12):1037-46.
PubMed 9872460 Garcia MF, Gordon MN, Hutton M, Lewis J, McGowan E, Dickey CA, Morgan D, Arendash GW. The retinal degeneration (rd) gene seriously impairs spatial cognitive perfor mance in normal and Alzheimer's transgenic mice. Neuroreport. 2004 Jan 19;15(1):73-7.
PubMed 15106834 Hyde LA, Crnic LS, Pollock A, Bickford PC. Motor learning in Ts65Dn mice, a model for Down syndrome. Dev Psychobiol. 2001 Jan;38(1):33-45.
PubMed 11150059 Hymson DL, Hynes MD. Evidence that ethanol-induced impairment of roto-rod performance is not mediated by opioid mechanisms. Prog Neuropsychopharmacol Biol Psychiat ry. 1982;6(2):159-65.
PubMed 6287531 Le Marec N, Lalonde R. Sensorimotor learning and retention during equilibrium tests in Purkinje cell degeneration mutant mice. Brain Res. 1997 Sep 12;768(1-2):310- 6.
PubMed 9369330 McFadyen MP, Kusek G, Bolivar VJ, Flaherty L. Differences among eight inbred strains of mice in motor ability and motor learning on a rotorod. Genes Brain Behav. 2 003 Aug;2(4):214-9.
PubMed 12953787 Morris R. Developments of a water-maze procedure for studying spatial learning in the rat. J Neurosci Methods. 1984 May;11(1):47-60.
PubMed 6471907 Paylor R, Baskall-Baldini L, Yuva L, Wehner JM. Developmental differences in place-learning performance between C57BL/6 and DBA/2 mice parallel the ontogeny of hip pocampal protein kinase C. Behav Neurosci. 1996 Dec;110(6):1415-25.
PubMed 8986342 Pompl PN, Mullan MJ, Bjugstad K, Arendash GW. Adaptation of the circular platform spatial memory task for mice: use in detecting cognitive impairment in the APP(SW ) transgenic mouse model for Alzheimer's disease. J Neurosci Methods. 1999 Feb 1;87(1):87-95.
PubMed 10065997 Prusky GT, West PW, Douglas RM. Behavioral assessment of visual acuity in mice and rats. Vision Res. 2000;40(16):2201-9.
PubMed 10878281 Schellinck HM, Forestell CA, LoLordo VM. A simple and reliable test of olfactory learning and memory in mice. Chem Senses. 2001 Jul;26(6):663-72.
PubMed 11473932 Schellinck, H. M., Forestell, C. A., & LoLordo, V. M. The development of a simple test of olfactory learning and memory. In: Marchlewska-Koj, A. Lepri, J. J., Mull er-Schwarze, D. Chemical Signals in Vertebrates 9 (book) (pp 459-465). Kluwer Academic/Plenum Publishers, 2001.
Wolfer DP, Stagljar-Bozicevic M, Errington ML, Lipp HP. Spatial Memory and Learning in Transgenic Mice: Fact or Artifact?. News Physiol Sci. 1998 Jun;13:118-123.
PubMed 11390774 Yerkes, R. M. The dancing mouse: A study in animal behavior. In: The animal behavior series. Vol 1. (89-113). MacMillan. 1907.